SFO-PCV Société Française d'Orchidophilie de Poitou-Charentes et Vendée

par J.P. RING*
à ROBERT… _ dont je garde le souvenir ému d’un esprit curieux et ouvert sur toutes les Choses de la Nature lui qui par la noblesse de son métier d’Agriculteur la côtoyait journellement dans sa rudesse et son âpreté. _ Toute une vie n’aura pas suffi à combler sa soif de connaissances. Du plus modeste caillou ramassé dans ses vignes aux myriades d’étoiles qui scintillent au firmament, tout était objet d’interrogation et d’émerveillement.

Cette année encore, tout comme les années précédentes la saison orchidologique démarre avec l’incontournable Ophrys sphegodes, toujours avec le même plaisir renouvelé et le même enthousiasme car il s’agit des premières retrouvailles sur le terrain après la longue trêve hivernale. _ Au début les premiers contacts sont empressés, histoire de retrouver leur mine changeante. Aussi on visite chaque pied en essayant de n’en vexer aucun par son oubli, ce qu’on n’a pas l’habitude de faire avec un Ophrys apifera ou un Ophrys scolopax pourtant très polymorphes eux aussi. _ Et puis les choses se gâtent progressivement. Devant le caractère déroutant de leur polymorphisme outrancier on finit par tourner la tête à leur approche comme pour leur signifier que leurs fantaisies agacent. Oui j’avoue, j’entretiens avec Ophrys sphegodes ce genre de rapports orageux.
Les moyens offerts par les technologies modernes peuvent se montrer redoutables pour espérer débusquer les secrets de cette remarquable variabilité. Voilà près de trente ans que je les poursuis de mes assiduités et ma collection de diapositives est garnie de plus d’une centaine de clichés de cette espèce, mais sans jamais avoir pu lever le moindre voile du mystère qui les entoure. _ Les possibilités offertes par le numérique devraient se montrer autrement plus efficaces. Grâce aux énormes capacités de mémoire de ces bijoux de technologie je vais enfin pouvoir les enfermer dans ma boîte magique pour leur faire livrer leur secret.
Ce sont aujourd’hui plus de mille clichés numérisés qui viennent s’aligner en rangs serrés dans les mémoires de mon ordinateur. Je les revisite de temps en temps surtout pendant les longues soirées d’hiver lorsque leur absence les fait de nouveau espérer, histoire aussi de les interroger une à une et de leur faire révéler leur secret : en vain. _ En effet lorsque vous arrivez à la trentième vous avez déjà oublié toutes les subtilités de l’organisation que vous offrait la première.
Aussi j’ai décidé de leur tirer à chacune un portrait robot sur papier que je découpais ensuite de façon à faire des regroupements selon un certain nombre de critères comme la forme de la macule, les rapports entre longueur et largeur de leur labelle, la taille des gibbosités…. _ Je me suis vite lassé devant la multiplicité des combinaisons et me suis résolu à l’étude d’un seul caractère, celui-là même qui m’intriguait le plus, à savoir les caractéristiques extrêmement changeantes de leur macule dans la perspective évidemment de vérifier si ce caractère pouvait être retenu dans une étude ultérieure sur la probable hétérogénéité des populations d’Ophrys sphegodes.
Je me suis donc remis devant mes planches de dessin, faisant fi des formes du labelle, de la taille des gibbosités, de la présence ou de l’absence d’un ourlet jaune, de l’écart entre les pseudos yeux…pour ne m’attacher qu’à ce seul caractère de la taille et de la forme très précise de la macule. Le document joint montre quelques unes des planches réalisées à ce jour soit plus de trois cents fleurons étudiés. (Planche I, Planche II, Planche III)

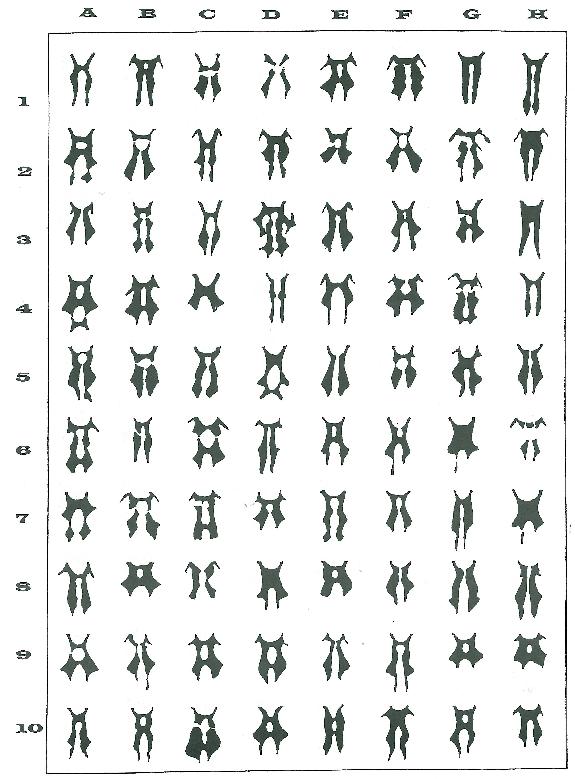

Les premiers dessins se faisaient plutôt en tirant la langue et le coup de crayon peu assuré cherchait maladroitement à respecter scrupuleusement les moindres contours à première vue sans grande cohérence d’une fleur à l’autre. Au fil des dessins le trait s’est brusquement accéléré comme si subitement il enjoignait le pas à une certaine logique. Rapidement les vagues pressentiments se sont mués en certitudes et il n’aura pas fallu plus de trente silhouettes dessinées pour qu’une lueur jaillisse et que comme l’inspecteur Bourrel je m’exclame subitement : « …mais oui ! mais bien sûr ! »
En effet il m’est vite apparu que les macules souvent si complexes et si diverses n’étaient que la répétition un nombre défini de fois d’un même motif de base.
Quelques exemples bien choisis mais parfaitement conformes à de multiples cas d’observation comme chacun pourra le vérifier sur le terrain tout comme à partir des silhouettes des planches dessinées (Planches I, II et III) sont tout à fait aptes à générer la même suggestion chez chacun.
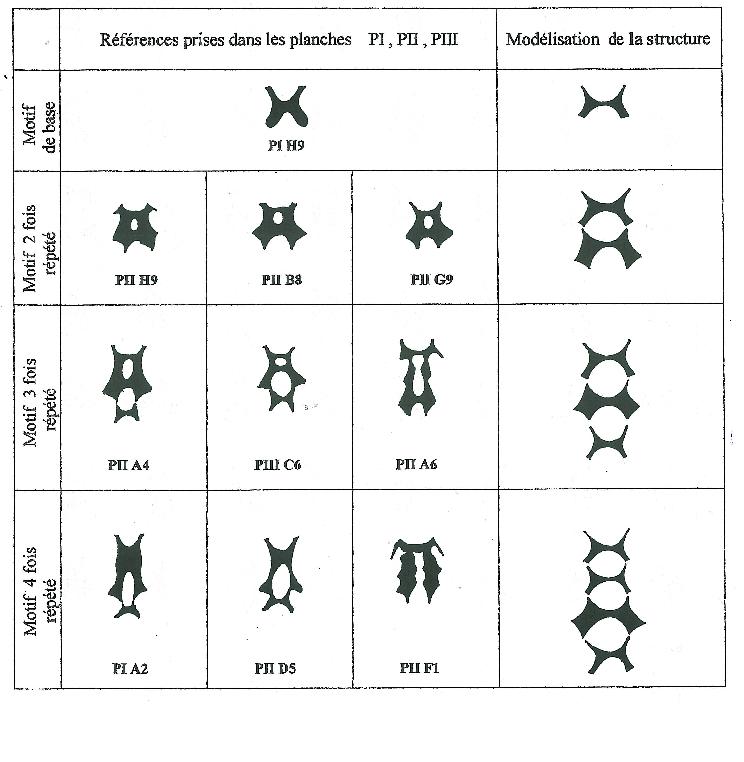
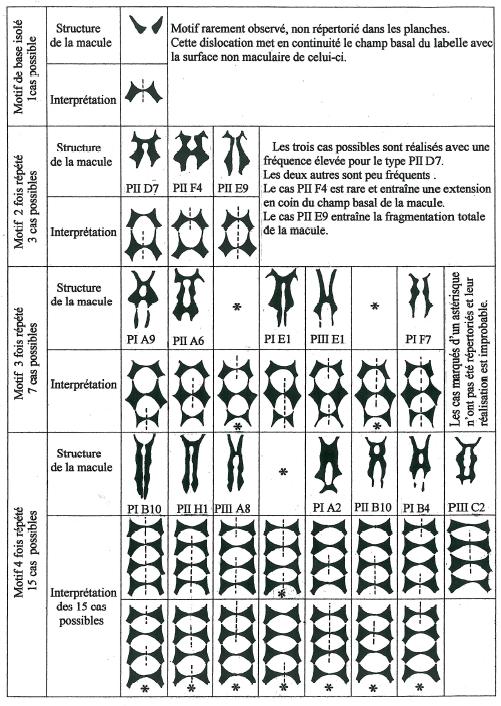
On remarquera aisément que plus le nombre de répétitions du motif de base est élevé et plus les structures deviennent incomplètes ou présentent des modifications.
Ainsi la « perforation » qui se situe à la limite de deux motifs consécutifs n’est pas toujours présente et de nombreuses macules « imperforées » et donc pleines doivent être interprétées comme répétitives, comme modélisé sur la figure 2. C’est ainsi qu’il convient d’interpréter les macules à motif quatre fois répété de la figure 1.
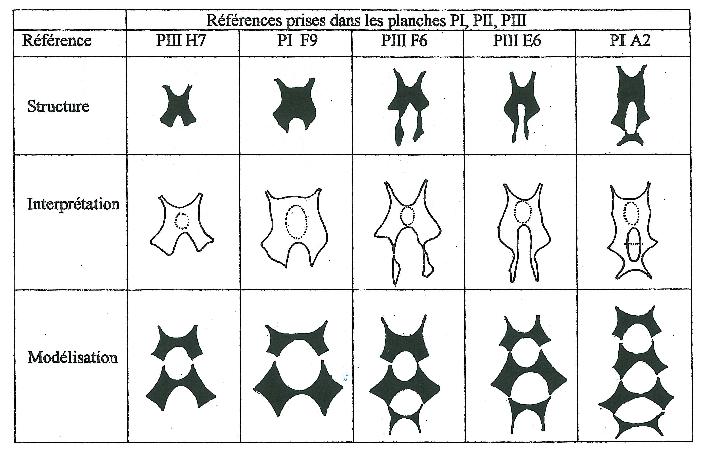
Perforation et imperforation ne sont pas à considérer comme des caractères individuels et coexistent sur le même pied sur des fleurons différents. La figure 3 représente les macules de sept fleurons distincts observés sur un même pied et le cas peut être généralisé.
Il n’y a donc pas à prendre en compte ce caractère dans une éventuelle discrimination entre individus, mais uniquement le nombre de répétitions du motif de base, caractère qui lui est parfaitement stable sur un pied déterminé et donc individuel et génétiquement programmé.
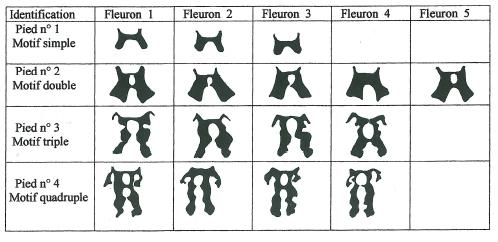
La figure 4 qui correspond au détail des macules de quatre pieds différents choisis pour leur représentativité des quatre groupes à un, deux, trois et quatre motifs montre clairement que le nombre de répétitions du motif de base et parfaitement constant pour un pied donné quelle que soit par ailleurs la complexité de cette macule.
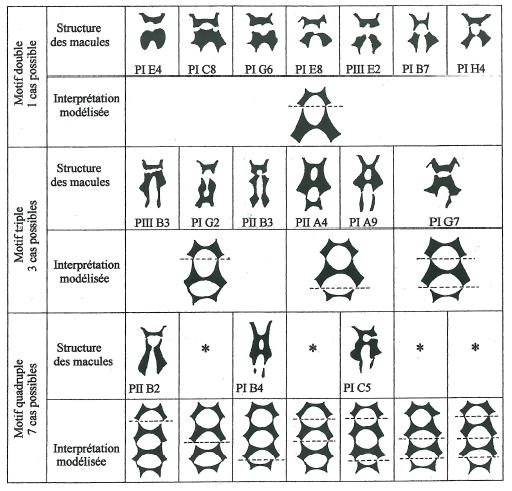
Parmi les autres modifications qui apparaissent avec la répétition des motifs de base la disparition du pont entre les structures collatérales est remarquable. Rare dans un motif de base isolé ou deux fois répété elle devient quasi systématique à partir des motifs triples et quadruples, pouvant même aller jusqu’à la scission complète de la macule dans le sens de la longueur. _ La figure 6 illustre cette possible fragmentation axiale de la macule en rapport avec le degré de répétition du motif de base. _ Une rapide étude statistique à partir des macules figurées sur les trois planches montre que quel que soit le cas de figure : motif de base isolé, deux, trois ou quatre fois répété, le motif de base proximal c’est à dire limitant le champ basal, est rarement fragmenté. (il reste entier dans près de 85 % des cas). _ Par contre cette fragmentation s’accentue pour l’échelon distal de la macule et d’autant plus que la répétition du motif de base est élevée. Elle affecte l’échelon distal de près de 95 % des macules à quatre motifs. (Figure 5)
Il ressort de ces observations que la macule bien que présentant une structure répétitive se caractérise par une forte asymétrie proximo-distale. C’est la raison pour laquelle la figure 6 fait apparaître des combinaisons modélisées absentes dans la réalité, non point par insuffisance d’échantillonnage ou par mauvaise interprétation des structures, mais simplement parce que la plupart des solutions symétriques de celles effectivement observées n’ont quasiment pas de chance d’être réalisées. _ Une sorte de portrait robot (figure 7) peut donc être tiré à présent, élaboré à partir des résultats statistiques sur la polarité proximo-distale de la macule et des combinaisons les plus fréquemment réalisées répertoriées dans le tableau de la figure 6.
Structures répétitives, forte polarité antéro-postérieure sont aussi les caractéristiques fondamentales de l’organisation d’un grand nombre d’organismes animaux et en particulier des insectes dont l’organisation segmentaire est flagrante..
Nous savons déjà que les Orchidées sont capables d’utiliser leur potentiel génétique pour réaliser la synthèse à l’identique des phéromones des insectes. D’ici à imaginer qu’elles utilisent de même leur potentiel génétique pour mimer le plan d’organisation de ces derniers n’a rien de délirant surtout lorsqu’on se réfère aux progrès récents réalisés dans le domaine de la génétique sur le développement.
Il ne suffit pas en effet que soit réalisé l’état pluricellulaire pour que soit constitué un organisme métazoaire ou métaphyte. Un important pool de gènes qualifiés de gènes du développement permettent aux insectes tout comme à nous mêmes de réaliser une organisation complexe en mettant en place des structures répétitives qu’on appelle des métamères ou segments dont la réalisation est gouvernée par les gènes de segmentation. Nous-mêmes conservons en nous les traces de cette segmentation sous la forme de vertèbres et de la musculature associée.
Chez les organismes évolués comme les insectes et nous mêmes la mise en place d’une polarité antéro-postérieure est sous la dépendance de gènes du développement qu’on appelle en langage savant les gènes homéotiques et qui permettent aux segments d’acquérir leur position fonctionnelle. C’est sous l’effet de tels gènes que le tagme céphalique portera des antennes et le tagme thoracique trois paires de pattes chez l’insecte.
La connaissance de ces gènes a surtout évolué grâce à l’observation de mutations comme la mutation « antennapedia » qui fait que l’insecte porte sur sa tête une paire de pattes en position et place d’une paire d’antennes. Une telle substitution porte le nom d’ homeosis. Certains considèrent que le fait de posséder des doigts surnuméraires ou des paires de mamelles en surnombre serait dû à des gènes homéotiques mutés et constitueraient donc un phénomène d’homéosis.
On voit donc apparaître une parfaite unité dans le monde vivant pour ce qui concerne les mécanismes de construction du plan d’organisation. C’est ainsi qu’on a trouvé chez la drosophile ( la petite mouche du vinaigre très étudiée en génétique par Morgan et son équipe) une séquence d’ADN commune avec le génome de la souris et que les généticiens appellent une homéoboîte ou homéobox. Plus récemment des séquences d’ADN similaires ont été identifiées dans le règne végétal et des gènes homéotiques sont bel et bien impliqués dans le développement des végétaux.
On voit donc qu’il n’est point irraisonnable de penser que les Orchidées possèdent le potentiel génétique pour réaliser une simulation de l’organisation segmentaire c’est à dire répétitive de leurs pollinisateurs.
Reste à comprendre comment ce potentiel génétique va être utilisé et exprimé pour parvenir à la réalisation des structures répétitives observées. Ce sera l’objet de cette deuxième partie basée comme la précédente sur les faits d’observation.
L’intervention de gènes homéotiques dans la mise en place des unités répétitives de la macule est purement spéculative de façon à … « faire avancer le Schmildblick »
Un certain nombre d’observations semblent néanmoins accréditer cette hypothèse.
Parmi celles-ci l’existence de dislocations transverses, déjà rencontrées dans certains exemples cités, et qui matérialisent en quelque sorte le caractère répétitif.
La figure 8 présente quelques unes des macules les plus représentatives choisies parmi celles des planches PI, PII, PIII.
On remarquera que les dislocations son fréquentes entre le premier et le deuxième échelon ainsi qu’entre le dernier et l’avant dernier, mais que dans le cas d’un motif quadruple celles ci ne tombent jamais entre les échelons intermédiaires. Les motifs intermédiaires forment de ce fait un bloc inséparable. _ Une hiérarchie proximo-distale semble ainsi se dessiner entre les différents motifs tout à fait caractéristique d’un déterminisme à caractère homéotique de l’agencement segmentaire, à l’image de ce que l’on observe chez les animaux et en particulier chez les insectes.. _ Cette hiérarchie pourrait être modélisée comme suit pour une macule à double, triple ou quadruple motif en se basant sur les fréquences les plus élevées des macules observées.
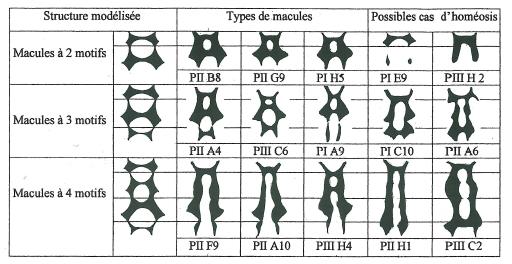
L’échelon proximal limitant le champ basal du labelle est de ce fait toujours limité dans son extension et se prolonge en collier autour de la colonne du gynostème.
Quel que soit le cas de figure, macule à deux, trois ou quatre échelons, le deuxième échelon a toujours une grande extension mais plus particulièrement dans les macules à deux et trois échelons .
Dans le cas des macules à quatre échelons c’est le troisième qui a généralement l’extension maximale.
Pour les macules à trois et quatre motifs l’échelon distal apparaît toujours très régressé, quasi atrophique voire complètement vestigial ou même à trace unilatérale.
Une étude statistique sur 500 fleurons a révélé que le type à trois échelons est largement prédominant et pourrait donc constituer le type de base dont on pourrait faire dériver les autres types.
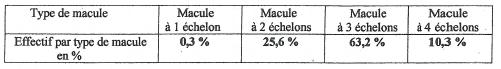
L’effectif de 25 % attribué aux macules à deux échelons est probablement même exagéré au détriment du type à quatre échelons étant donnée la difficulté dans certains cas à faire la discrimination avec le type à trois échelons du fait de la forme en pointe du motif distal, pointe pouvant parfois correspondre à un troisième motif vestigial.
Différents mécanismes génétiques gouvernés par des gènes de segmentation et des gènes homéotiques pourraient être à la base de la variation autour du type à trois motifs que nous considérerons comme le type fondamental.
La figure 10 donne une vision synoptique des filiations possibles entre les différent types de macules ainsi que des mécanismes génétiques qui sont à la base de leur réalisation.
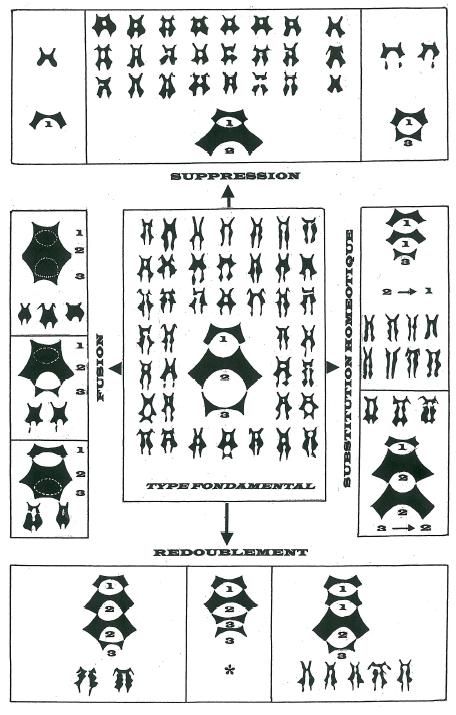
Quatre grands mécanismes génétiques paraissent être à la base de la plupart des variations autour du type fondamental à trois motifs. _ Le mécanisme de suppression assure le passage à des macules à deux ou exceptionnellement à un motif. _ Le type 1-2 peut s’expliquer par la suppression de l’échelon 3 par mutation conservée dans les populations à une fréquence plus ou moins élevée d’où sa relative abondance. _ Le type 1-3 nettement plus rare peut s’expliquer par la suppression de l’échelon 2. _ Le type 1 limité à l’unique motif proximal implique une double mutation à l’origine de la disparition simultanée ou consécutive des motifs 2 et 3. Une double mutation est un évènement rare d’où la fréquence extrêmement faible de ce type dans les populations. _ A l’inverse du mécanisme de suppression le mécanisme de redoublement permet d’accrémenter d’une unité le nombre de motifs de la macule en particulier de faire passer le type fondamental de trois motifs à quatre. Trois cas sont théoriquement possibles. _ Le type 1-1-2-3 par redoublement de 1 est nettement le plus fréquent. _ Le type 1-2-2-3 par redoublement de 2 apparaît occasionnellement. _ Le type 1-2-3-3 n’apparaît pas dans mes échantillonnages et n’aurait de toute façon qu’ un intérêt mineur vu le caractère très vestigial du motif 3. _ Les mécanismes de substitution comme leur nom l’indique correspondent au remplacement d’un motif donné par un autre de position hiérarchique différente. Le fait de substituer un motif par un autre comme on le voit pour la mutation antennapedia chez la Drosophile où une paire de pattes vient prendre la place des antennes, correspond à un phénomène d’homéosis et est donc imputable à la mutation d’un gène homéotique. _ La substitution 2→ 1 c’est à dire remplacement d’un motif 2 par un motif 1 est le cas le plus fréquent et conduit à des macules à branches grêles particulièrement effilées très à part de l’aspect des autres macules et contribue fortement au caractère polymorphe et si déroutant des populations d’Ophrys sphegodes. Plusieurs mécanismes peuvent d’ailleurs être à l’origine de telles macules à branches grêles et de longueur très variable soit isolément, soit par action conjointe comme le montre le tableau de la figure 11.
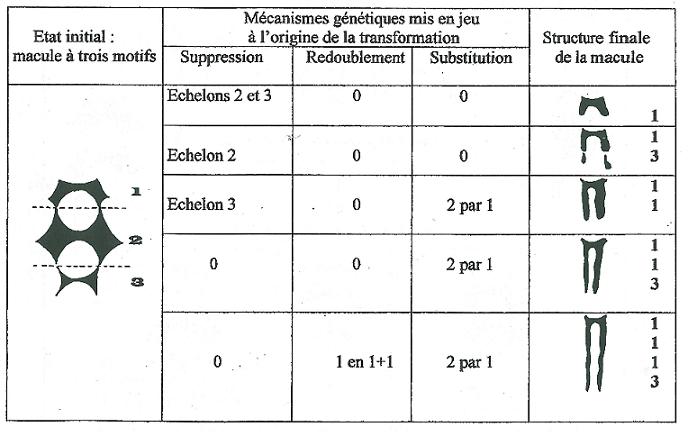
Contrairement à la substitution homéotique qui constitue le mécanisme majeur qui mène à la réalisation de macules grêles le mécanisme de fusion est toujours à l’origine de macules denses et trapues tendant vers une forme rectangulaire parfois parfaitement réalisée. La figure 10 montre les trois cas de fusion possible à partir du type fondamental. En aucun cas ces macules ne doivent être confondues avec les macules pleines « imperforées » citées plus haut, leur allure très caractéristique les en distingue nettement.
Les quatre mécanismes génétiques évoqués permettent une interprétation raisonnée de la grande majorité des macules rencontrées comme en témoigne l’échantillonnage non exhaustif présenté sur la figure 10 qui ne tient pas compte des mécanismes de fissuration longitudinale et transverse, partielle ou totale. _ Or ces images de dislocation en particulier celles où la fissuration longitudinale est couplée avec les coupures transverses sont particulièrement instructives sur les mécanismes qui sont à la base de la réalisation des macules et peuvent nous renseigner sur l’expression des gènes évoqués. _ La figure 12 donne en répertoire les macules affectées par de telles dislocations.

La dislocation totale est peu fréquente mais non rare, ici six cas de dislocation totale nette sur l’effectif des 280 macules figurées sur les trois planches. _ Le terme même de dislocation implique dans notre esprit que les structures observées résultent de la fragmentation d’une structure unique en éléments disjoints. _ Dans cette hypothèse les éléments éclatés devraient conserver les caractéristiques géométriques de la structure d’origine et la juxtaposition de quelques macules à différents degrés de dislocation donnerait une assez bonne image de la succession des évènements.(Figure 13)
Même si la représentation de la figure 13 permet d’interpréter les images de fissuration de la macule déjà en place, elle ne permet pas de donner un cadre explicatif à sa formation. _ L’entrée chronologique du tableau de la figure 13 pourrait alors tout aussi bien se faire par la droite pour aller de structures disjointes vers une structure de macule pleinement constituée.. _ Il ne s’agirait plus dans ce cas d’une dislocation, mais d’une convergence et d’une coalescence de motifs à l’origine éloignés. L’observation des images de la figure 13 suggèrerait alors que les zones rapprochées de ces motifs subiraient une attraction et un développement favorisé aussi bien dans le sens transversal que longitudinal du labelle et que les zones éloignées seraient freinées dans leur développement. _ Les macules qui en résulteraient présenteraient effectivement un aspect approchant de la réalité, et pourraient parfaitement expliquer les cas de dislocation transverse et longitudinale, mais présenteraient des contours peu conformes et surtout conduiraient vers des images de perforation quadrangulaires de la macule.( Figure 14) _ Mise à part la macule PII F4 (voir Planche II) cette situation n’est que très rarement réalisée.
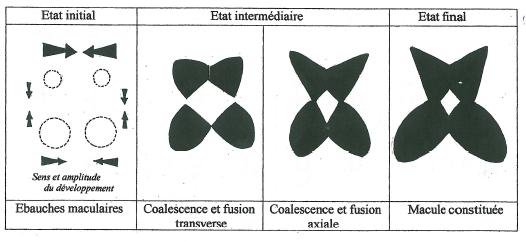
La distorsion entre la forme réelle de la macule à contours anguleux et notre modèle à formes beaucoup trop molles ainsi que la non conformité des perforations qui devraient tendre vers une structure circulaire à ovale nous amène à envisager l’intervention des structures externes à la macule. Il pourrait s’agir de « centres extramaculaires » inhibiteurs qui viendraient contrarier et moduler le développement des ébauches maculaires.
Cette hypothèse est évidemment basée sur l’observation d’un certain nombre d’anomalies sur lesquelles nous reviendrons.
La figure 15 développe un modèle très simple à la base de la structure d’une macule à deux motifs réalisée à partir du développement simultané et synchrone des ébauches maculaires et des centres inhibiteurs extramaculaires.
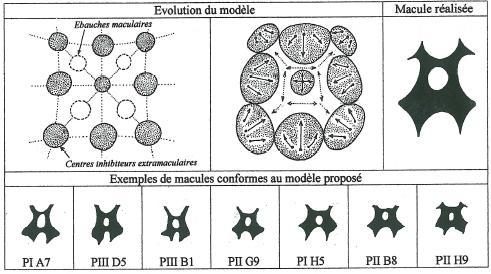
La conformité avec la réalité est flagrante. Nous voyons donc que la réalisation de la macule résulte autant du développement des centres extramaculaires que de celui des ébauches maculaires.
Ainsi la macule ne fait qu’occuper la surface du labelle non entamée par les centres extramaculaires, un peu à la façon d’un bas-relief.
Un équilibre dont va dépendre la forme de la macule doit donc s’établir entre le développement des ébauches maculaires et celui des centres inhibiteurs extramaculaires.
Si la balance penche en faveur des ébauches maculaires celle-ci va acquérir des formes molles et des perforations maculaires quadrangulaires en conformité avec le modèle de la figure 14.
Si au contraire elle penche en faveur des centres extramaculaires nous nous orientons vers des macules anguleuses à limites concaves et à perforations rondes ou ovales en conformité avec le modèle de la figure 15.
La figure 16 donne les résultats du fonctionnement à l’équilibre ou en déséquilibre des deux types de structures dans le cadre de la construction d’une macule à deux motifs.
Nous voici devant des perspectives beaucoup plus complexes que la simple prise en compte de la structure répétitive de la macule, c’est l’organisation du labelle tout entier et sa régionalisation qu’il nous faut envisager à présent, et en particulier sa zone basale, trop complexe au départ mais dont nous pouvons entreprendre l’étude maintenant.

Dans toute l’étude qui précède nous avons toujours occulté la complexité de la région proximale de la macule dont nous savons qu’elle embrasse la colonne du gynostème. Généralement filiforme elle peut occasionnellement présenter deux élargissements qui correspondent à deux autres paires d’ébauches maculaires associées à des centres extramaculaires.
Il est par ailleurs très fréquent que la face interne des gibbosités prenne le même aspect que la macule elle même et se montre en continuité avec elle, ce qui correspond au développement d’une autre paire latérale d’ébauches maculaires qui réalisent la fusion avec les ébauches contiguës du même rang.
Toutes ces caractéristiques sont portées dans la figure 17 qui retrace la géographie de l’ensemble des ébauches maculaires ainsi que des centres extramaculaires associés.
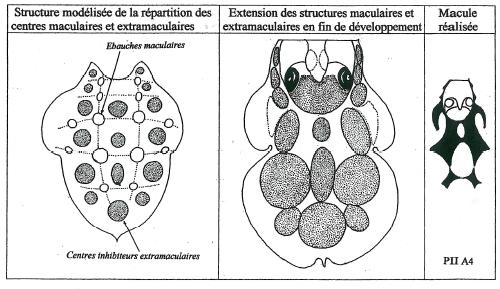
La parfaite concordance entre la structure maculaire issue du fonctionnement du modèle et la structure réelle des macules observables est remarquable et nous incite à valider notre modèle qui paraît nettement plus complexe que ne le laissait présager notre première approche. _ La parfaite alternance des bandes maculaires avec les centres extramaculaires confirme l’organisation répétitive de la structure des macules qui imitent de façon tout à fait remarquable l’organisation segmentaire du corps des insectes. Chez ces derniers chaque segment en formation est déterminé en demi segments antérieur et postérieur interagissant par des mécanismes inducteurs et inhibiteurs. On pourrait tout aussi bien considérer la structure du labelle comme une succession d’unités segmentaires, six exactement, chacune constituée d’un rang d’ébauches maculaires associée à un rang de centres extramaculaires en interaction activatrice et inhibitrice. _ Mais les similitudes s’arrêtent là. On ne peut en effet assimiler le complexe mécanisme organogène de la mise en place de l’ensemble des organes de l’insecte au contrôle histogène probablement réalisé sur le seul tissu épidermique à l’intérieur d’un organe déjà déterminé dans sa structure et qui est le labelle. _ Une étude histologique du labelle au cours de son développement devrait nous éclairer sur le bien fondé et la crédibilité de notre modèle en mettant en évidence les éventuelles actions activatrices et inhibitrices au niveau des structures épidermiques.
Les coupes histologiques de la figure 18 donnent un aperçu de la structure du labelle d’un fleuron épanoui d’Ophrys sphegodes et permettent de comprendre les caractères tinctoriaux et de structure des zones maculaires et extramaculaires.
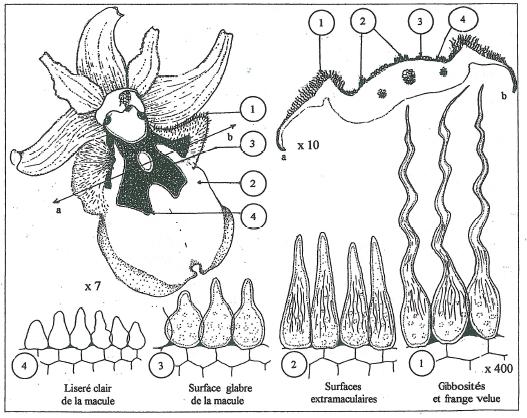
Les coupes réalisées montrent très nettement l’implication des cellules épidermiques de la face supérieure du labelle dans les caractéristiques structurales ( pilosité des gibbosités, velouté des surfaces non maculaires, glabrescence de la macule ) et tinctoriales (couleur brune à brun-rouge des surfaces non maculaires, aspect brun clair très changeant de la macule, présence d’un discret liseré jaunâtre à crème limitant la macule.) _ Aussi ne prendrons nous en compte que les caractéristiques histologiques de cet épiderme à l’exclusion de tout autre tissu du labelle. _ Quatre principaux types cellulaires peuvent être distingués. _ Le type (1) de la figure 18 est caractéristique des gibbosités et de la frange pileuse en périhérie du labelle et détermine leur aspect fortement velu. _ Les cellules sont différenciées en longs poils unicellulaires de forme ondulante et à vacuole chargée d’anthocyanes à l’origine de la couleur brun-rouge de ce trichome. _ Le type (2) recouvre toute la surface non maculaire du labelle à l’exclusion du liseré jaune qui borde certains labelles. L’aspect joliment velouté de cette surface est due à des cellules coniques en forme de massue et à vacuole également remplie d’anthocyanes. L’aspect finement cannelé de la paroi cellulaire tout comme de celle du type cellulaire précédent peut être à l’origine du caractère chatoyant des couleurs. _ Le type (3) en forme d’outres trapues recouvre toutes les surfaces de type maculaire. _ Le caractère très ramassé et bas des cellules est responsable de l’aspect glabre de la macule. _ La charge en anthocyanes des vacuoles détermine pareillement la coloration mais d’intensité différente par rapport aux surfaces précédentes pour des raisons que nous évoquerons ultérieurement. _ Enfin le type (4) caractérise l’étroite frange de couleur jaunâtre à crème qui limite le champ maculaire. L’observation attentive rien qu’à la loupe révèle la constance de ce liseré qui peut parfois être très discret. L’absence d’anthocyanes et la forme très ramassée des cellules à aspect simplement papillaires, parfois presque isodiamétriques sont à l’origine de l’aspect caractéristique de cette zone. _ Il apparaît donc clairement que l’épiderme de la face supérieure du labelle est un tissu hautement différencié qui au delà de son rôle protecteur exprime le potentiel génétique qui l’adapte à sa fonction de simulation des insectes pollinisateurs. _ La juxtaposition des quatre types cellulaires dans l’ordre de la figure 18 incite évidemment à envisager les étapes de cette différenciation au cours du développement. _ Le type (4) est rapidement arrêté dans son développement alors que le type (1) subit le développement le plus poussé tout en passant par les étapes intermédiaires (2) et (3).
Pour la réalisation de cette étude j’ai disséqué des boutons floraux d’âge différent dont certains très précoces et dont j’ai effectué des coupes. _ Certains points d’observation se sont révélés très instructifs pour notre problématique. Les principaux sont consignés dans la figure 19. _ Au début du développe alors que le labelle a une forme simplement ovale sans trace de gibbosités celui-ci a un aspect foliaire à nervation très caractéristique délimitant des espaces parenchymateux, donnant à l’ensemble du labelle une belle coloration verte d’un beau vert-fluo. _ A ce stade les cellules épidermiques sont très basses, simplement bombées sans autre trace de pigmentation que la pigmentation chlorophyllienne. Elles sont donc très peu différenciées. On ne distingue à ce stade pas la moindre trace de macule, ni de champ basal. Bien que les gibbosités soient encore absentes les cellules épidermiques de leur territoire présomptif ont commencé leur différenciation en prenant un aspect plus allongé. _ On voit ensuite le labelle progressivement s’épaissir, les gibbosités démarrer leur développement par le creusement de deux fossettes remontant par la face inférieure. _ De verte, la pigmentation vire sensiblement et de façon très diffuse vers le brunâtre en commençant par la périphérie du labelle dans la zone des gibbosités ainsi que la frange velue au niveau des centres extramaculaires. La macule commence discrètement à se dessiner tout en conservant sa couleur verte.
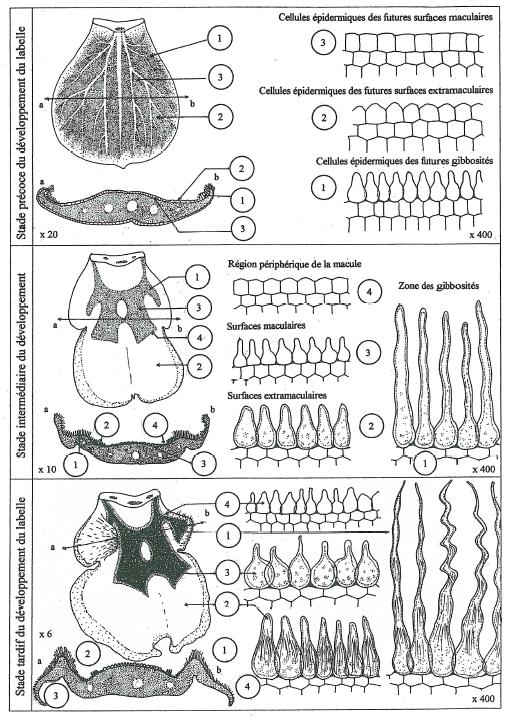
Les cellules des gibbosités sont à présent nettement allongées, leur vacuole est colorée d’un beau rouge limpide par les anthocyanes. C’est la superposition de la pigmentation chlorophyllienne à localisation chloroplastique avec la pigmentation anthocyanique d’origine vacuolaire qui est à l’origine de la coloration brune à l’échelle macroscopique _ La figure 19 montre la succession des transformations des cellules épidermiques au fil du développement jusqu’au stade final du bouton floral prêt à l’éclosion.
La succession des évènements tant au niveau de l’évolution des types cellulaires que de celui de la pigmentation vient corroborer de façon attendue ou tout à fait inattendue tant la démonstration est flagrante, notre modèle de réalisation de la macule.
En effet on voit les cellules épidermiques se différencier en commençant par les centres extramaculaires parfaitement dessinés, faisant ainsi apparaître par contraste le dessin de la macule d’abord dépourvue de toute pigmentation anthocyanique et donc verte. Les cellules des centres maculaires restent momentanément inhibés dans leur développement par les zones extramaculaires jusqu’à un moment tardif où elles se soustraient partiellement de cette inhibition pour prendre leur forme et leur pigmentation caractéristique.
Seules les cellules à la frontière des ébauches maculaires et des centres inhibiteurs subissent une inhibition prolongée et quasi totale, formant ainsi le fin liseré jaunâtre qui encercle la macule. Cette zone d’inhibition totale est particulièrement étroite chez Ophrys sphegodes, d’où la finesse du liseré.
Chez d’autres Ophrys comme apifera ou scolopax cette inhibition reste beaucoup plus puissante, entraînant ainsi la formation d’un large liseré jaune autour de la macule. Seules les cellules de la macule à distance des centres inhibiteurs subissent une différenciation, incomplète et tardive faisant apparaître juste avant l’éclosion du bouton floral une pigmentation intramaculaire brune mais très différente de la pigmentation extramaculaire : mélange chagriné de plages brunes et de plages jaunâtres traduisant l’inhibition exercée. (Figure 20)
Chacun pourra revenir à sa collection de diapo. sur ces espèces ou à des ouvrages de haute qualité d’illustration comme celui de la Société Française d’Orchidophilie (Les Orchidées de France, Belgique et Luxembourg Collection Parthénope, pages 288 à 223) et ainsi regarder avec un œil nouveau ces belles macules si esthétiquement cerclées de jaune.
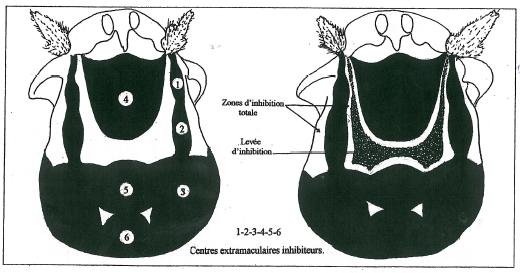
Tout mécanisme biologique aussi parfait soit-il a ses avatars et ses ratés. Il en va de même de celui que nous venons d’établir. Loin de contredire notre modèle et de l’invalider ces erreurs ou faux pas de la nature ne font que le confirmer et le renforcer.
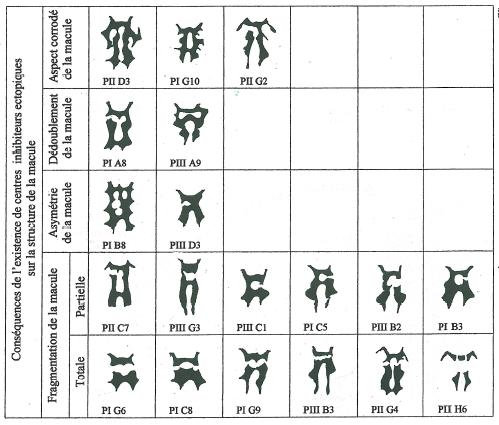
Ainsi l’existence de macules à aspect corrodé comme celles figurées sur la figure 21 sont la confirmation éclatante de l’existence de centres inhibiteurs extramaculaires. Mais au lieu de présenter la parfaite disposition comme représenté sur la figure 17, ces centres peuvent être morcelés et en position ectopique, c’est à dire à une place qui en principe n’est pas la leur. _ L’existence de tels centres inhibiteurs ectopiques a plusieurs conséquences sur la structure de la macule des pieds qui les possèdent comme en témoignent les images de la figure 21. _ Elle peut d’abord donner comme on vient de le préciser un aspect corrodé à la macule, d’autant plus que le morcellement de ces centres est important. _ Elle peut ensuite entraîner un dédoublement de la structure de la macule, le plus fréquemment dans la région proximale près du champ basal. _ L’apparition encore plus anarchique de tels centres ectopiques peut rendre asymétrique la structure de la macule. _ Mais surtout, et c’est le cas très nettement le plus fréquent l’existence de tels centres est à l’origine de multiples cas de fragmentation de la macule déjà décrits par ailleurs, fragmentation transverse, totale ou partielle. La fragmentation partielle est évidemment génératrice d’asymétrie.
L’existence de labelles hypochromes, autre particularité parfois rencontrée, est le résultat d’une mutation affectant la chaîne de synthèse des pigment anthocyaniques vacuolaires. Cette anomalie jette un éclairage particulier sur le mécanisme de différenciation des cellules épidermiques du labelle. Elle montre en effet de façon très claire la disjonction qui existe entre les caractères structuraux des cellules et les caractères tinctoriaux. _ La non synthèse des anthocyanes n’interfère en rien avec la synthèse des pigments chlorophylliens qui est correctement réalisée dans les chloroplastes, ce qui confère au labelle une coloration verte. _ Cette forme d’hypochromie est aussi qualifiée de viridisme. _ L’existence et la forme de la macule n’en sont pas pour autant affectés, ce qui montre bien qu’ au cours de la différenciation cellulaire la réalisation de la structure des cellules épidermiques est totalement indépendante de l’acquisition de la pigmentation.
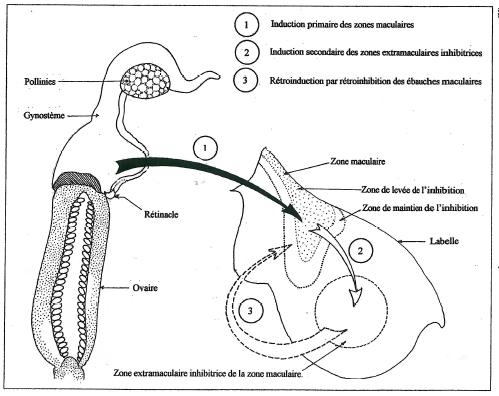
Parmi les lusus couramment rencontrés chez les Ophrys celui de la labellisation des pétales latéraux peut présenter un intérêt majeur pour élucider l’enchaînement des mécanismes d’activation-inhibition qui sont à l’origine de la macule et en particulier les mécanismes inducteurs qui initient l’ensemble du processus. _ J’ai en effet souvent remarqué que ce type de lusus s’accompagnait d’anomalies et de malformations au niveau du gynostème. Il semblerait donc que l’ensemble du processus qui aboutit à la construction du labelle et à sa régionalisation ait comme point de départ les organes fertiles de la fleur, ce qui nous incite pour clore cette étude à proposer le schéma corrélationnel de la figure 22 fruit de l’ensemble des observations que j’ai pu effectuer à ce jour sur les Ophrys.
En conclusion à cette étude disons qu’elle est l’aboutissement d’une longue période d’errements. _ Comme beaucoup sans doute j’ai eu avec les Ophrys mais plus particulièrement avec Ophrys sphegodes de très sérieuses difficultés face au polymorphisme des populations. _ Les tentatives d’études statistiques visant à démontrer une éventuelle hétérogénéité des populations et en particulier à évaluer leur degré d’introgression par Ophrys araneola se sont toujours soldées par un échec chaque fois qu’il s’agissait de prendre en compte les caractéristiques de la macule, critère à mon avis majeur voire incontournable pour une telle étude, et ceci à cause de l’incapacité à trouver des paramètres quantifiables et non pas simplement descriptifs car dérivant alors vers une classification vite incontrôlable vue l’avalanche des cas particuliers.
Cette étude a eu pour premier objectif celui de me doter d’un outil fiable de mesure des variations observables au niveau des motifs aussi variés que ceux de la macule. _ Il est vrai que cet outil fonctionne et qu’aucun cas de macule aussi complexe soit-il ne peut plus échapper à l’analyse. Le modèle proposé étant susceptible de trouver un cadre explicatif à n’importe quel cas de figure. C’est ce qui me conforte pour valider le modèle proposé. _ Il est vrai aussi que je me pose beaucoup de questions face à la pertinence ou à l’impertinence d’une telle publication. C’est évidemment l’avantage et le suprême intérêt d’une revue comme la nôtre de pouvoir exposer ses idées en toute liberté et sans prétention aucune. L’extrême diversité des nombreux articles présentés en témoignent. Jamais en effet je ne me serais permis une telle publication dans une revue de grande diffusion, tant je suis critique moi-même sur ma propre démarche. Personne à ce jour n’a à ma connaissance prospecté dans cette direction des gènes du développement pour trouver un cadre explicatif au polymorphisme labellaire. _ D’autres mécanismes génétiques comme ceux de la variabilité somatique pourraient tout aussi bien servir d’hypothèse.
Seul face à mes angoisses je serais évidemment heureux de recevoir l’avis des lecteurs en retour, en particulier après avoir testé le modèle proposé comme je le suggère dans l’article. Il est toujours frustrant suite à une publication de ne pas recevoir d’écho positif ou négatif. Je n’imagine pas en effet que tous ceux qui ont éprouvé les mêmes _ difficultés n’y trouvent pas un quelconque intérêt. Tout au moins certains regarderont-ils dorénavant le labelle des Ophrys avec un regard nouveau. Rien qu’en cela l’article aura fait œuvre utile en exerçant chez chacun une vision plus critique et moins routinière.
L’objectif premier étant atteint à savoir la dotation d’un outil de mesure de la diversité maculaire, nous allons pouvoir l’utiliser à présent. _ Dans un premier temps pour voir si les paramètres définis sur l’organisation de la macule sont suffisamment discriminants pour distinguer les espèces très proches d’Ophrys sphegodes, plus pariculièrement O.araneola, mais aussi O.Argensonensis et quelques autres types à petit labelle pour le moment non classés. _ On pourra ensuite envisager l’étude de populations suspectées hétérogènes, mesurer le degré d’introgression des populations d’O. sphegodes par O. araneola par exemple.
Enfin il serait particulièrement intéressant de confronter « le modèle sphegodes » avec d’autres taxons en particulier avec les fucifloridés, les diverses formes d’apifera… _ Essayez car vous verrez qu’avec Ophrys sphegodes nous nous sommes attaqués au cas le plus coriace et le plus récalcitrant. Interpréter une macule d’un scolopax ou d’un apifera devient à côté un jeu d’enfant auquel je vous invite vivement à vous adonner : c’est passionnant.
* 1333, route des Bruères - 86550 Mignaloux-Beauvoir